
Изучение филогенетических взаимоотношений игуаноморфных ящериц представляет собой ключевую проблему для понимания происхождения, родственных связей и эволюционной истории распространения чешуйчатых рептилий. Основная после Кэмпа (Camp, 1923) концепция филогении ящериц, высказанная Сухановым (1961) и получившая свое развитие в исследованиях Эстеса (Estes, 1983, 1985) рассматривает игуаноморфных ящериц (Iguania), объединяющих три семейства: Agamidae, Chamaeleonidae и Iguanidae как монофилетическую группу, сестринскую по отношению ко всем остальным чешуйчатым.
Об общем происхождении игуаноморфных ящериц говорят факты, полученные при изучении строения и развития зубной системы в онтогенезе разных семейств чешуйчатых. У всех игуаноморфных ящериц, имеющих, как известно, зубную систему с принципиально разным порядком замещения и прикрепления зубов (плевродонтная у игуанид и акродонтная у агамид и хамелеонов), был обнаружен особый тип закладки и строения яйцевых зубов. Яйцевой зуб — это характерное только для чешуйчатых рептилий эмбриональное приспособление для вскрытия твердой оболочки яйца. Проведенное нами сравнительное изучение эмбрионов показало, что у всех современных ящериц и змей яйцевые зубы имеют два типа закладки — парную и непарную. Парную закладку имеют все Gekkota, Scincomorpha, Anguinomorpha и все змеи (Ophidia), в то время как всех Iguanomorpha объединяет синапоморфия — непарное состояние закладки яйцевых зубов (Ананьева, Орлов, 1986).
Акродонтные ящерицы — группа (надсемейство), объединяющая два семейства — Agamidae и Chamaeleonidae, и признанная монофилетической практически всеми исследователями, как следует из названия, характеризуются акродонтным строением зубов, то есть их прикреплением к верхнему краю челюстной кости. Их объединяет также отсутствие плоскости разлома внутри позвонков, по которой происходит интервертебральная автотомия хвоста у большинства других ящериц. Относящиеся к этому надсемейству ящерицы распространены, в основном, на территориях гондванального происхождения (Мадагаскар, Сейшельские острова, Африка, Аравийский полуостров, Индия, Юго-Восточная Азия и Австралия), а также в тех районах Азии, которые имеют лавразийский генезис.
Разнообразное по внешнему облику и приспособлениям к различным условиям существования семейство Agamidae объединяет свыше 50 родов и около 350 видов ящериц крупных, средних или мелких размеров (длина туловища с хвостом от 8 см у мелких круглоголовок до 1 м у парусной ящерицы) с дневной активностью. Семейство включает наземные, древесные, роющие формы, а также виды, способные к планирующему полету (род Draco) и водному образу жизни (роды Hydrosaurus и Physignathus). Важнейший признак, отличающий агам от большинства других ящериц — акроплевродонтное и гетеродонтное состояние их зубной системы.
Важным этапом в развитии филогенетических взглядов на эту группу чешуйчатых следует считать исследование Муди (Moody, 1980), посвященное изучению филогенетических и биогеографических взаимоотношений родов семейства Agamidae, где было использовано 122 морфологических признака, относящихся к строению скелета, мускулатуры и покровов. В результате этой работы была предложена филогенетическая система агамовых ящериц, выделены 6 основных групп, обоснована самостоятельность ряда родов, до того времени рассматривавшихся в составе более крупных. Так, например, в составе рода Agama была восстановлена самостоятельность пяти разных родов. Предложенные в 1980 году Муди представления о филогенетических взаимоотношениях внутри семейства не утратили своей актуальности до настоящего времени и во многом нашли подтверждение в последующих исследованиях по молекулярной филогении.
В составе семейства Муди выделяет следующие группы: группа I включает крупных примитивных растительноядных наземных ящериц родов Leiolepis и Uromastyx; группа II — крупных примитивных растительноядных ящериц родов Hydrosaurus и Physignathus, ведущих водный образ жизни; группа III — целый ряд родов наземных ящериц, эволюционировавших в Австралии; группа IV — меланезийский и австралийский род Hypsilurus. Основная часть современных родов агамовых ящериц отнесена этим автором к группам V (разнообразные древесные агамы южной и юго-восточной Азии) и VI (наземные и петрофильные агамы аридных районов Африки и западной Азии).
В работе были использованы материалы (более 1000 экземпляров) из систематических коллекций Зоологического института РАН, Зоологического Музея Московского государственного университета, Зоологических Музеев США (Музей Естественной истории Филда, Калифорнийская Академия Наук, Музей Зоологии Позвоночных Калифорнийского Университета Беркли, Национальный музей естественной истории Смитсониан), Австралии (Австралийский музей), Канады (Королевский Музей Онтарио), Германии (Зоологический Музей им А. Кенига, Гамбургский Зоологический музей, Зоологический Музей Университета им. А. Гумбольдта, Дрезденский Музей природоведения), Франции (Музей естественной истории), Китая (Институт Биологии Ченгду, Китайская Академия Наук) и Японии (Тропический Биосферный Научный центр, Университет Рю Кю).
Для исследования покровов гистологическими методами и с использованием растровой электронной микроскопии были взяты образцы из разных частей головы, спинной и брюшной стороны тела и хвоста с последующей соответствующей обработкой. При растровом электронно-микроскопическом исследовании использовались увеличения от 50 до 2000.
Анализ митохондриального генома (Macey et al., 2000) показал, что внутри монотипической группы акродонтных ящериц выделяются семь клад. Интересно отметить довольно четкое «географическое» и «экологическое» представительство выделенных эволюционных линий. Они представлены следующими группами: 1) Chamaeleo, 2) Uromastyx, 3) Leiolepis, 4) Hydrosaurus, 5) юго-восточно-азиатская группа видов, 6) афро-западноазиатская группа видов и, наконец, 7) австралийско-новогвинейская группа, в которую входит также обитающий в юго-восточной Азии вид Physignatus cocincinus. Самостоятельность и монофилия всех выделенных групп подтверждается с высокой долей статистической вероятности. Различные монотипические группы акродонтных ящериц проникли в Азию, вероятно, разными путями. Две клады, уромастицины и западноазиатско-африканская клада, попали туда либо через Индию, либо через Африку; юго-восточноазиатская клада — либо через Индию, либо через Юго-Восточную Азию. Очевидное юго-восточноазиатское происхождение имеют три группы: лейолеписы, гидрозавры и физигнатусы. Таким образом, прослеживается различная эволюционная история основных групп акродонтных ящериц, связанная со сложной тектонической эволюцией массивов, на которые распалась Гондвана. Доказательства привлекаются из данных анализа ДНК: наиболее глубокая филогенетическая дивергенция внутри каждой клады указывает на ее древнее происхождение.
Построенные филогенетические деревья показывают, что хамелеоны представляют собой сестринскую группу по отношению ко всем агамовым ящерицам, а также дают основания для разделения семейства агамовых ящериц на несколько подсемейств, которые соответствуют эволюционным линиям, выделение которых подтверждено результатами анализа митохондриальной ДНК. Внутри семейства агамовых ящериц выделяются шесть подсемейств: Uromastycinae Theobald, 1868 c видами рода Uromastyx, Leiolepidinae Fitzinger, 1843 с видами рода Leiolepis; Amphibolurinae Wagler, 1830 со всеми австралийскими и новогвинейскими видами, а также Physignathus cocincinus; Hydrosaurinae Kaup, 1828 с видами рода Hydrosaurus; южноазиатское и юго-восточноазиатское подсемейство Draconidae Fitzinger, 1826 с многочисленными индийскими и юго-восточно-азиатскими родами агам, ведущих преимущественно древесный образ жизни, характеризующееся максимальным богатством родов; и, наконец, афро-западноазиатское подсемейство Agaminae Spix, 1825. Последнее представляет собой афро-азиатскую ветвь радиации семейства, представленное наземнообитающими пустынными и горными ящерицами, количество родов которых далеко не столь разнообразно, как в южной и юго-восточной Азии (Ананьева, 2004).
За последнее десятилетие достигнут значительный прогресс в изучении таксономического разнообразия и филогенетических отношений внутри отдельных родов подсемейства Draconinae, что, несомненно, важно для понимания состава, происхождения и филогеографических трендов азиатской фауны агамовых ящериц (Honda et al., 2000; Macey et al., 2000; Ананьева, 2004). Проведены ревизии родов Acanthosaura (Kalyabina-Hauf et al., 2004; Ананьева и др., 2006; Orlov et al., 2006), Bronchocela (Hallermann, 2004, 2005), Calotes (Ota, Hikida, 1991, 1996; Hallermann, 2000; Vindum et al., 2003; Zug et al., 2006); Draco (Inger, 1983; Musters, 1983; McGuire, Heand, 2001), Gonocephalus (Honda et al., 2002), Japalura (Ota, 1989, 1991, 2000; Ota et al., 1998), Pseudocalotes (Hallermann, Bohme, 2000; Hallermann, McGuire, 2001), Ptyctolaemus (Manthey, Nabhitabhata, 1991; Ananjeva, Stuart, 2001; Schulte et al., 2004). В ходе этих ревизий, среди прочих результатов, были сделаны описания ряда новых видов.
Исследования митохондриального генома (Macey et al., 2000) и морфологических особенностей покровов (Ananjeva et al., 2001) свидетельствуют о монофилии клады, представляющей виды рода Leiolepis и о самостоятельности этой эволюционной ветви, ранее объединяемой с родом Uromastyx в одно подсемейство (или даже отдельное семейство) Uromastycinae Theobald, 1868). Эта монотипическая группа в своем происхождении связана с юго-восточноазиатскими плитами, ведущими свое начало от Гондваны. Представители подсемейства отличаются развитием бедренных пор и рецепторами без волоска (Ananjeva, 1997, Ананьева, 2004). Среди видов известны как бисексуальные виды (L. belliana, L. guttata, L. reevesii, L. peguensis), так и партеногенетические, триплоидные (L. triploida, L. guentherpetersi) или диплоидные (L. boehmei) (Darevsky, Kupriyanova, 1993).
Подсемейство Draconinae — наиболее представительная группа по количеству родов древесных и полудревесных агам. Среди них как монотипические и представленные одним — тремя видами роды (Aphaniotis, Cophotis, Ptyctolaemus, Sitana), так и обширные, с высоким видовым разнообразием (Draco, Gonocephalus, Japalura). Для этой эволюционной линии акродонтных ящериц характерен высокий процент эндемичных родов. Муди (Moody, 1980) отмечает 14 эндемичных родов на Индийском субконтиненте и 15 эндемичных родов в Юго-Восточной Азии, что вместе составляет более чем половину всех родов семейства (53 рода, согласно Муди). Столь высокая диверсификация на родовом уровне — отличительная черта подсемейства, филогенетические взаимоотношения внутри которого долгое время оставались неясными. Исследования последних лет позволяют расширить наши представления о составе и филогении подсемейства (рис. 1), хотя остается немало спорных моментов даже в отнесении вновь описанных или повторно исследованных видов к тому или иному роду (Manthey, Grossmann, 1997; Macey et al., 2000; Ananjeva, Stuart, 2001; Schulte et al., 2004). Результаты анализа митохондиальной ДНК (Macey et al., 2000) свидетельствуют о полифилии рода Japalura.
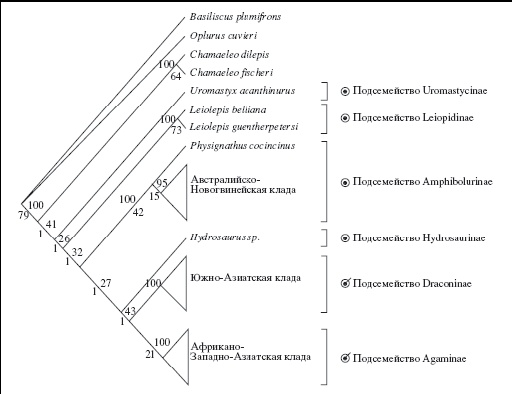
Рис. 1
Проблемы филогенетических взаимоотношений родов внутри подсемейства Agaminae вызывают большой интерес исследователей. Для исследователей фауны Палеарктики особенно актуальны исследования родов Laudakia, Trapelus, Phrynocephalus c высоким таксономическим разнообразием и их родственных взаимоотношений. Данные анализа митохондриальной ДНК (Macey et al., 2000, 2006) позволяют утверждать, что род Laudakia представлен двумя самостоятельными эволюционными линиями. Род Trapelus представляет собой сестринскую группу по отношению к виду Bufoniceps laungwalensis, первоначально описанному в составе рода Phrynocephalus. Уже получив ранг самостоятельного рода, Bufoniceps рассматривался как сестринский таксон по отношению к роду Phrynocephalus (рис. 2). Эта точка зрения в настоящее время опровергнута. Внутри рода Trapelus на основании данных анализа митохондриального генома подтверждена видовая самостоятельность видов Trapelus agilis и T. sanguinolentus (Macey et al., 2000, 2006).
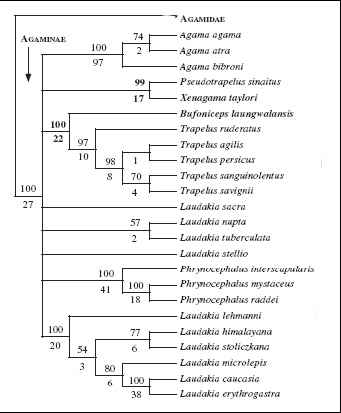
Рис. 2
Попытка оценить приуроченность покровных образований с эволюционными линиями внутри семейства показала, что один тип кожных органов чувств (рецептор с волоском) типичен только для 2 подсемейств: Draconinae и Agaminae, а у остальных агам развиты линзоообразные рецепторы. Агамовые ящерицы демонстрируют высокое морфологическое разнообразие эпидермальных голокринных желез. Бедренные поры характерны для агам подсемейств Uromastycinae, Leiolepidinae, Hydrosaurinae, Amphibolurinae (кроме Chelosania и Moloch) и рода Mantheyus, принадлежащего к подсемейству Draconinae (Ананьева, Дуйсебаева, в печати). Бедренные поры не развиты у агам подсемейств Agaminae и Draconinae (за исключением рода Mantheyus). У агам родов Agama, Trapelus, Acanthocercus и Laudakia (подсемейство Agaminae) развиты уникальные морфологические структуры: эпидермальные голокринные железы (Dujsebayeva, 1998).
Следует отметить, что применение современных молекулярно-генетических методов не только позволяет оценить и уточнить представления о филогении основных групп агамовых ящериц и их таксономическом разнообразии, но и дает новый импульс для изучения морфологических признаков и выявления наиболее перспективных направлений таксономических исследований.
Н. Б. Ананьева
Зоологический институт РАН, Санкт-Петербург
Ананьева Н. Б., 2004. Филогения и биогеография агамовых ящериц (Agamidae, Lacertilia, Reptilia): обзор концепций и результатов молекулярных и морфологических исследований // Успехи cовременной биологии. Т. 124. № 1. С. 44—57.
Ананьева Н. Б., Дуйсебаева Т. Н., 2007. Бедренные поры агамовых ящериц (Agamidae, Sauria, Reptilia) // Современная герпетология. Т. 7. В печати.
Ананьева Н. Б., Орлов Н. Л., 1986. Строение и развитие яйцевых зубов чешуйчатых рептилий и некоторые аспекты их филогении // Труды ЗИН АН СССР. Л. Т. 157. С. 34—41.
Ананьева Н. Б., Орлов Н. Л., Калябина-Хауф С. А., 2006. Криптическое таксономическое разнообразие тропических агамовых ящериц Acanthosaura, Agamidae, Sauria) // Успехи современной биологии. Т. 126. № 5. С. 505—513.
Суханов В. Б., Некоторые вопросы филогении и системы Lacertilia (s. Sauria) // Зоол. ж. 1961. Т. 40. Вып. 1. С. 73—83.
Ananjeva N. B., 1997. Diversity of agamids (Sauria, Reptilia): Ecological forms and morphology
of skin receptors // In: H. Ulrich (ed.). Tropical biodiversity and systematics. Proc. Intern. Symposium on Biodiversity and Systematics in Tropical Ecosystems.
Bonn, 1994. Zoologisches Forschungsinstitute und Museum Alexander Koenig.
Bonn. S. 297—304.
Ananjeva N. B., Dujsebayeva T. N., Joger U., 2001. Morpological study of squamate
integument: more evidence for the metataxon status of Leiolepidinae // Journ.
Herpetol. V. 35. № 3. P. 69—74.
Ananjeva N. B., Stuart B., 2001. The agamid lizard Ptyctolaemus phuwuanensis Manthey
and Nabhitabhata, 1991 from Thailand and Laos represents a new genus // Russian
Journal of Herpetology. V. 8. № 3. P. 165—170.
Camp C. L., 1923. Classifi cation of the lizards // Bull. Amer. Mus. Nat.Hist. V. 48. Art. 11.
P. 289—481.
Darevsky I. S., Kupriyanova L. A., 1993. The two all-female lizard liecies of the genus
Leiolepis Cuvier, 1829 from Thailand and Vietnam (Squamata: Sauria: Uromastycidae)
// Herpetozoa. Bd. 6. Hf. 1/2. S. 3—20.
Dujsebayeva T. N., 1998. The histology of callous scales of the males of Asian rock
agamas, Laudakia caucasia and Laudakia himalayana (Reptilia: Agamidae) //
Russian Journal of Herpetoljgy. V. 15. № 2. P. 160—164
Estes R., 1983. Handbuch der Paleoherpetologie. Teil 10/A. Sauria terrestria. Amphisbaenia.
Gustav Fischer Verlag. Stuttgart — New York. 245 pp.
Estes R., 1985. Phylogenetic relationships and biogeography of squamates // In: Z. Rocek
(ed.). Abstracts Third Ordin. Gen. Meet. SEH. Prague. P. 60—61.
Hallermann J., 2000. A new species of Calotes from the Moluccas (Indonesia) with notes
on the biogeography of the genus (Sauria: Agamidae) // Bonn. Zool. Beitr. Bd. 49.
Hf. 1—4. S. 155—163.
Hallermann J., 2004. A new species of the genus Bronchocela from the tropical rain
forest of southern Vietnam // Russian Journal of Herpetoljgy. Moscow. V. 11. № 1.
P. 30—34.
Hallermann H., 2005. A taxonomic review of the genus Bronchocela (Squamata:
Agamidae) with description of a new species from Vietnam // Russian Journal of
Herpetology. V. 12. № 3. P. 167—182.
Hallermann J., Böhme W., 2000. A review of the genus Pseudocalotes (Squamata:
Agamidae) with description of a new species from West Malaysia //
Amphibia — Reptilia. 21: P. 193—210.
Hallermann J., McQuire J. A., 2001. A new species of Pseudocalotes (Squamata:
Agamidae) from Bukit Larut, West Malaysia // Herpetologica. V. 57. № 3.
P. 255—265.
Honda M., Ota H., Kobayashi M., Nabhitanhata J., Hoi-Sen Yong, Sengoku S., Hikida T.,
2000. Phylogenetic relationships of the family Agamidae (Reptilia: Iguania) inferred
from mitochondrial DNA sequences // Zool. Sci. V. 17. № 5. P. 527—537.
Honda M., Ota H., Sengoku S., Hoi-Sen Yong, Hikida T., 2002. Molecular evaluation
of phylogenetic signifi cance in the highly divergent karyotypes of the genus
Gonocephalus (Reptilia: Agamidae) from tropical Asia // Zool. Sci. V. 19. № 1.
P. 129—133.
Inger R. F., 1983. Morphological and ecological variation in the fl ying lizards (genus
Draco) // Fieldiana Zoology. New Ser. № 18. Publ. 1346. P. 1—37.
Kalyabina-Hauf S., Ananjeva N., Joger U., Lenk P., Murphy R.W., Stuart B. L., Orlov
N. L., Cuc Tho Ho, Wink M., 2004. Molecular phylogeny of the genus Acanthosaura
(Agamidae) // Current Herpetol. V. 23. № 1. P. 7—16.
Macey J. R., Schulte J. A., II, Larson A., Ananjeva N., Wang Y., Pethiyagoda R., Rastegar-
Pouyani N., Papenfuss T., 2000. Evaluating Trans-Tethys Migration: An Example
Using Acrodont Lizard Phylogenetics. Syst. Biol. V. 49. № 2. P. 233—256.
Macey J. R., Schulte J. A., Fong J. J., Das I., Papenfuss T., 2006. The complete mitochondrial
genome of an agamid lizards form the Afro-Asian subfamily Agaminae and the
phylogenetic position of Bufoniceps and Xenagama. // Molecular Phylogenetics and
Evolution. V. 39. P. 881—886.
Manthey U., Nabhitabhata J., 1991. An agamid lizard, Ptyctolaemus phuwuanensis Manthey
& Nabhitabhata 1991 (Sauria, Agamidae) from Northeast-Thailand // Sauria
(E). Kempton Park. Bd. 1. № 2. P. 3—6.
Manthey U., Grossmann W., 1997. Amphibien and Reptilien Südostasien. Natur and Tier
Verlag. Münster. 512 pp.
McGuire J. A., Kiew Bong Heand., 2001. Phylogenetic systematics of Southeast Asian
fl ying lizards (Iguania: Agamidae: Draco) as inferred from mitochondrial DNA sequence
data // Biol. Journal of the Linn. Soc. V. 72. P. 203—229.
Moody S., Phylogenetic and historical biogeographic relationships of the genera in the
family Agamidae (Reptilia, Lacertilia). 1980. Unpublished Ph. D. Thesis. Univ.
Musters C. J. M., 1983. Taxonomy of the genus Draco L. (Agamidae, Lacertilia, Reptilia)
// Zool. Verhandelingen. V. 199. P. 1—120.
Orlov N. L., Nguyen Quang Truong, Nguyen Van Sang, 2006. A new Acanthosaura allied
to A. capra Guenther, 1861 (Agamidae, Sauria) from Central Vietnam and Southern
Laos // Russian Journal of Herpetology. V. 13. № 1. P. 61—76.
Ota H., 1989. A new species of Japalura (Agamidae: Lacertidae: Reptlia) from Taiwan //
Copeia. V. 45. № 3. P. 569—576.
Ota H., 1991. Taxonomic redefi nition of Japalura swinhonis Günther (Agamidae: Squamata),
with a description of a new subspecies of J. polygonata from Taiwan // Herpetologica.
V. 47. № 3. P. 280—294.
Ota H., 2000. Japalura szechwanensis, a junior synonym of H. fasciata // Journ. Herpetol.
V. 34. № 4. P. 611—614.
Ota H., Hikida Ts., 1991. Taxonomic review of the lizards of the genus Calotes Cuvier
1817 (Agamidae: Squamata) from Sabah, Malaysia. // Tropical Zoology. V. 4.
P. 179—192.
Ota H., Hikida Ts., 1996. The second specimen of Calotes kinabaluensis de Grijs (Squamata,
Agamidae) from Sabah, Malaysia, with comments on the taxonomic status of
species // J. Herpetol. V. 30. № 2. P. 288—291.
Ota H., Szu-Lund Chen, Gaus Shang, 1998. Japalura luei: A new agamid lizard from
Taiwan (Reptilia: Squamata) // Copeia. V. 54. №. 3. P. 649—656.
Schulte J. A., II, Vindum J., Htun Win, Thin Thin, Kyi Soe Lwin, Awab Khwi Shein., 2004.
Phylogenetic relationships of the genus Ptyctolaemus (Squamata: Agamidae), with a
description of a new species from the Chin Hills of western Myanmar // Proc. Calif.
Acad. Sc.. Ser. 4. V. 55. № 12. P. 227—247.
Vindum J. V., Htun Win, Thin Thin, Kyi Soe Lwin, Awan Khein, Kla Tun., 2003. A new
Calotes (Squamata: Agamidae) from the Indo-Burman Range of western Myanmar
(Burma) // Proc. Calif. Acad. Sc. Ser. 4. V. 54. № 1. P. 1—16.
Zug G. R., Brown H. H. K., Schulte J. A. II, Vindum J. V., 2006. Systematics of the Garden
Lizards, Calotes versicolor Group (Reptilia, Squamata, Agamidae), in Myanmar:
Central Dry Zone Populations // Proc. Calif. Acad. Sc. Ser. 4. V. 57. № 1.
P. 35—68.
